

2014年の私達のOsCERK1の論文が出てすぐに、イタリアのMycファクターの存在を2007年に示したグループから接触がありました。彼らは2013年にAM菌の共生シグナルはNodファクターにそっくりなMyc-LCOと呼ばれる分子ではなく、キチンの短い分子である可能性を報告しています。彼らはその受容体がOsCERK1である可能性を考えて、oscerk1変異体を使って共生の初期応答であるカルシウムスパイキングを観察したいとの非常に丁寧なメールを送ってきたのです。
実は私達は既にその実験を計画していて、必要な遺伝子組換え植物を作って予備的なデータなどを得ていたのですが、私はポスドクの身分に過ぎなかったこともあって戦力が足りないことに加えて、別のプロジェクトで忙しくなっていたこともあり、彼らに共同研究に入っていただき、ついでに主導権も渡しました。結果として予想通りoscerk1変異体では短いキチンに対するカルシウムスパイキングが破綻しており、私が確認していた遺伝子発現よりも早い応答の起動にもOsCERK1が関わっていることが示されました。ちなみにこの論文は、New Phytologist (IF=7.43)という雑誌に掲載されています(私が発表したPCPよりずっと評価が高い)。
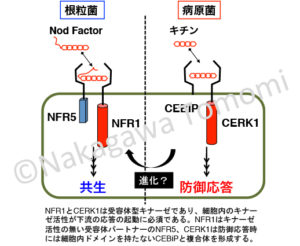
NFR1の場合は、同じくLysM型受容体キナーゼであるNFR5と受容体複合体を形成して共生応答を起動します。NFR5のキナーゼドメインは活性が無く、主に細胞外ドメインがNodファクターの識別に関わっていると考えられていますが、キナーゼとして以外の役割を果たしている可能性は否定できません。そこで根粒菌共生においてNFR5のキナーゼドメインが必須かどうかを調べたところ、下図のように重要であることが判りました。しかもイネのOsNFR5のキナーゼドメインでも代用できます。
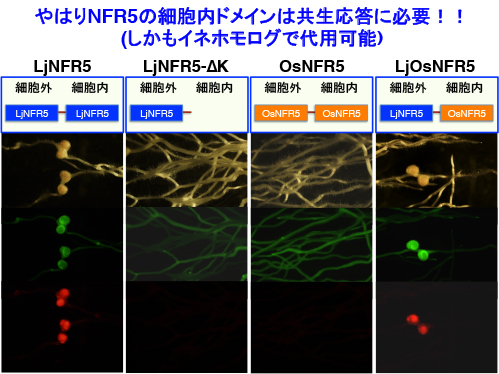
そこでOsNFR5の完全破壊変異体を作出したのですが、なんと全く正常に共生してしまいました・・・。上述のイタリアのグループとの共同研究でも、osnfr5変異体は正常なカルシウムスパイキングが観察されるので、どうやら共生と関係無いようです。したがってCERK1の共生応答時における受容体パートナーの有無などは、現状では全く見当もつきません。
コメント