
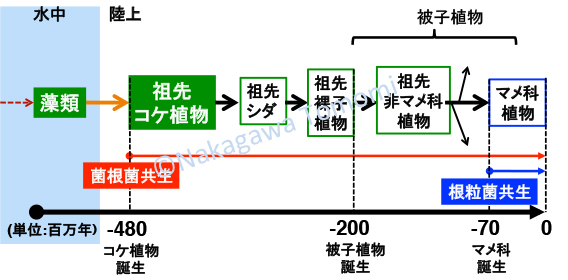
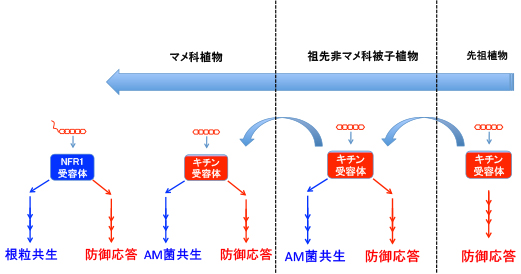
2011年にシロイヌナズナのAtCERK1とミヤコグサのNFR1の比較で、共生応答の誘導に重要なYAQ配列を同定しました (植物の敵と味方 〜承〜)。しかしマメ科植物の根粒菌共生に関わると思われたこの配列は、実はAM菌共生する大部分の植物のCERK1に保存されており、YAQや類似したYAR配列を持たないアブラナ科のCERK1は例外であることが判明しました(植物の敵と味方 〜転〜 1)。私はYAQ/YAR配列がアブラナ科以外のAM菌共生する被子植物のCERK1に保存されていた時点で、これは進化の話になると直感しました。なぜならばAM菌共生は、最初の陸上植物である原始のコケ植物で成立したと考えられるからです。
その後にCERK1は少なくともイネで二重機能を持っていることを私達は明確に証明しましたが(植物の敵と味方 〜転〜 3)、最初から二重機能だったとは考えにくいです。最初は共生か防御かのどちらかが先に在って、後から二つ目の機能が獲得されたと考えるのが自然でしょう。渋谷先生とは、どちらが先にあったかについて、よく議論になりました。先生は防御が当然先にあっただろうと主張されますが、陸上植物の誕生前に病原菌が居たかどうかは不明です。私は案外、先に共生があったというオチもあると考えていました。どちらにせよ、2人で盛り上がるくらいの興味があるネタではあります。
もちろん先祖の植物を調べる手段は無いのですが、現存のコケ植物を解析すれば、ヒントが得られる可能性が高い。そこでコケのCERK1の機能を調べたいと考えました。
コケのモデル植物としては、ヒメツリガネゴケとゼニゴケが知られています。どちらも非常に研究しやすい植物なのですが、これらの植物もシロイヌナズナと同じようにAM菌共生しない「例外的な」植物でした。したがって新たなAM菌共生するモデルコケ植物が必要です。そこで私が注目したのが、モデルとして整備されているゼニゴケ(Marchantia polymorpha)に近縁のフタバネゼニゴケ(Marchantia paleacea subsp. diptera)です。そこで2011年頃から、フタバネゼニゴケのCERK1がAM菌共生や防御応答のどちらに機能しているかを調べるプロジェクトを開始しました。

コメント