
みなさんは、自分が運の良い方だと思いますか?
私はその点は全く自信がありません。
ボスや渋谷先生から提示されたのは、「ミヤコグサのNFR1以外のLysM型受容体をRNAiで発現抑制して共生への影響を調べる」でしたが、これはRNAiで変化が観察されて初めて意味があるようなアプローチで、当たりを引ける気がしませんでした。
(何も考えずに適当にRNAiをやっても、当たる人は当てます)
そこで私は、運が悪くてもハズレが無いアプローチとして、ミヤコグサの根で共生シグナルであるNod ファクターの応答と、防御応答を誘導するキチンやflg22の応答を比較する戦略を採りました。マイクロアレイという手法を使えば、ミヤコグサの大部分の遺伝子の変動を見ることができるので、少なくとも部分的には共通するものがあるだろうし、それを元に議論して論文にできるだろうという筋書きです。
最初にミヤコグサの根におけるキチンとflg22の応答を比較したところ、大部分が重なることが判りました。誘導される遺伝子はflg22の方が多いのですが、これは実際の根の様子を観察していてもflg22の方が強烈なので、納得できます。誘導される遺伝子のメンバーを見ても、典型的な基礎的防御応答の遺伝子群です。これにより、当時は誰も解析していなかったミヤコグサの基礎的防御応答遺伝子群を特定しました。

次にミヤコグサ根粒菌のNod ファクターで誘導される遺伝子群を同定しました。そして、それらの遺伝子群と上述の基礎的防御応答遺伝子群と比較して、下記のようにNodファクター特異的、キチンと共通、flg22と共通、キチンとflg22の両方と共通という具合に分類しました。すると予想外なことに、Nodファクターを投与した直後に誘導される遺伝子の大部分は、キチンやflg22と共通の防御応答遺伝子が大部分であることが判りました。
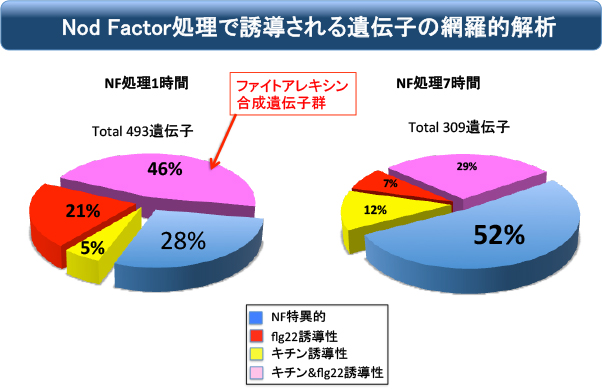
Nodファクターにだけ誘導されるのは約3割しか無く、7割は防御応答遺伝子です。したがって、「Nodファクター受容体は防御応答を引き起こす能力がある」という可能性が示唆されました。ちなみに7時間後にはNodファクター特異的な遺伝子が半分まで増えているので、おそらくは防御応答はすぐにキャンセルされて共生応答が主流になるようです。
もともとはNodファクターとキチンは非常によく似た物質です。キチン受容体がNodファクターを間違えて認識しただけかもしれません(自分にとってイヤな突っ込みをするのが重要)。これらの応答が本当にNFR1によって起動されたのかを確認するために、Real-Time PCRという方法で確認してみました。
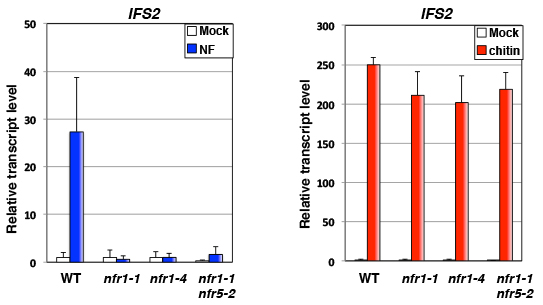
その結果、正常な野性型ミヤコグサ(WT)ではNodファクター(NF)によって抗菌物質の合成酵素であるIFS2が誘導されますが、NFR1が壊れたnfr1-1とnfr1-4の変異体や、NFR1とNFR5(NFR1のパートナー)の両方が壊れた二重変異体では全く誘導されなくなります。一方でキチンを投与した場合は、野性型も変異体も正常にIFS2が誘導されます。IFS2以外にも様々な防御応答遺伝子をチェックしましたが、傾向は全く同じでした。したがって
- Nodファクター投与による防御応答は、真にNFR1によって起動される。
- NFR1とは別に、ミヤコグサはキチン受容体を持っている。
ということが証明されました。
コメント