
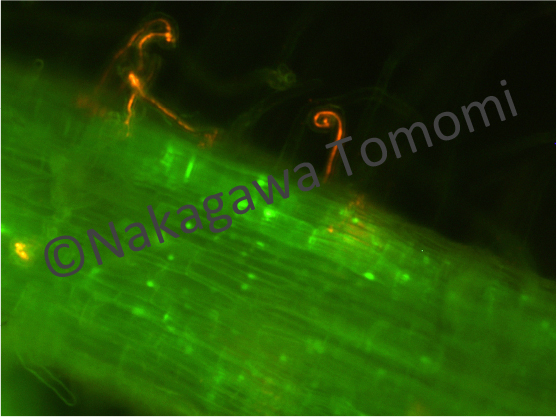
植物のLysM型受容体キナーゼは、2003年に同定されたマメ科植物のNFR1が最初だと思います。NFR1の果たす役割は絶大で、ミヤコグサのnfr1変異体はパートナーの根粒菌に対してほとんど応答しなくなってしまい、根粒菌は全く受け入れられません。つまりNFR1は根粒菌共生の開始を司る必須の受容体なのですが、植物業界で2007年に同定されたシロイヌナズナのCERK1受容体は、なんとカビの細胞壁成分のキチンを認識して排除する応答を起動するものでした。
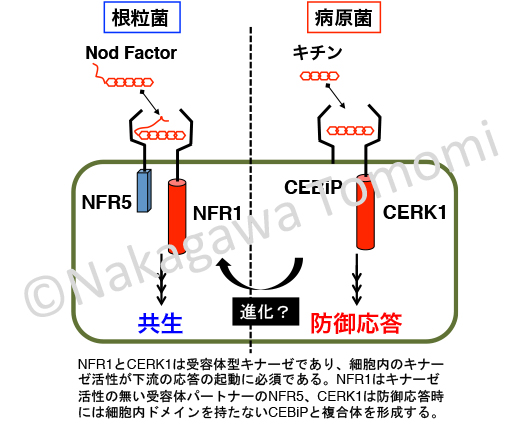
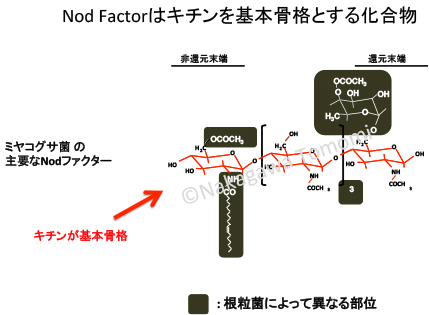
Nodファクターとキチンは非常に類似した物質なので、当然かもしれないですが、それぞれの生理応答を考えると非常に不思議です。
ミヤコグサのNFR1のアミノ酸配列でシロイヌナズナゲノムを検索すると、断トツで高い相同性を示すのがCERK1です。逆にCERK1でミヤコグサやタルウマゴヤシなどのマメ科モデル植物のゲノムを検索すると、NFR1を含めて多数がヒットしますが、どうやらNFR1/CERK1型の受容体はマメ科植物で大幅に数が増えたようです。また、アミノ酸配列の相同性だけではなく、それぞれの遺伝子の周辺の遺伝子の並び(microsynteny)も似ているという論文も出ていました。つまり共通の先祖遺伝子から進化した可能性が高いのですが・・・、生理的な応答が微生物の受け入れと排除という正反対なのです。
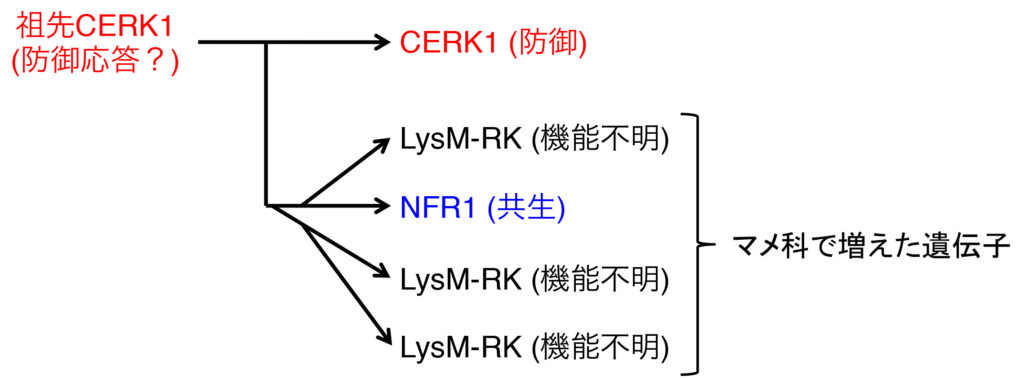
ちょうど私はミヤコグサで共生と防御応答の関係を調べて試行錯誤していた時期だったのですが、ボスがCERK1を発見した渋谷先生と仲が良かったので、「ミヤコグサにもNFR1とは別にCERK1があると思うから、その機能を調べてみないか?」というお話を頂きました。
コメント