
CERK1がAM菌共生に関わっている可能性が浮かび上がって来ました。これを確認するためには、CERK1が壊れた植物でAM菌共生を調べるのが王道です。シロイヌナズナではCERK1が壊れた変異体を渋谷先生達が単離していたのですが、残念ながらナズナはAM菌共生しません。
2010年に渋谷先生のグループが、イネ(Oryza sativa)のOsCERK1の発現をRNAiで低下させると、キチン防御応答が破綻することを報告しました。イネはAM菌が共生するので、私達はこのOsCERK1-RNAiの株を使わせてもらって徹底的に実験しましたが、微妙にAM菌共生が低下している気がするものの、統計的に有意な結果は得られませんでした。RNAiだとmRNAが半分から10%くらいまで低下しますが、それで充分な影響が出るかはケースバイケースです。私達は諦めきれずに試行錯誤していたのですが、渋谷先生の共同研究者である生物資源研究所(NIAS)の西澤洋子博士のグループが、ついにOsCERK1を完全破壊したoscerk1変異体を作出して下さいました。

西澤博士らのグループは、作出したoscerk1変異体の葉において、キチン(GN7)に対する活性酸素の発生や遺伝子発現が破綻していることを明らかにしています。我々が注目する根における応答についても調べるために、根の遺伝子発現の変化を次世代シーケンサーを使って解析しましたが、上図のようにoscerk1ではキチンに対して発現が上昇する遺伝子(緑)も低下する遺伝子(赤)も、応答しなくなっていることが確認できました。次にAM菌共生を調べました。その結果、oscerk1では見事にAM菌共生が止まっていたのです。期待はしていたのですが、実際にこの結果を見た時は震えました。
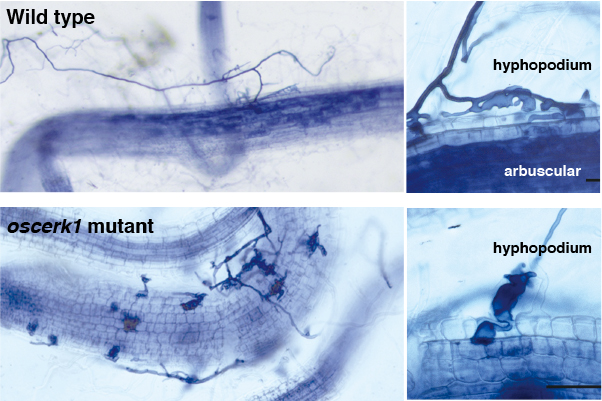
ちなみに西澤博士は、イネのキチン防御応答においてOsCERK1と受容体複合体を形成するOsCEBiPの変異体も作出していました。これを使わせて頂いて同様に実験したところ・・・
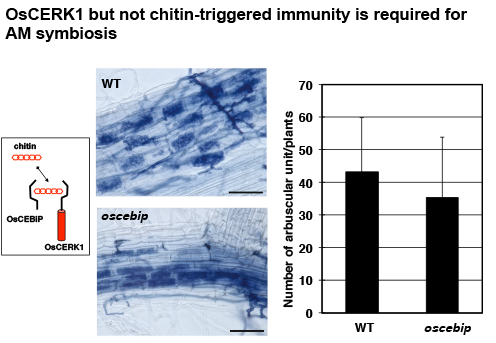
面白いことにこちらは全く影響がありません。つまりOsCERK1が大切なのであって、キチン防御応答自体はAM菌共生に関与していないのです。以上の結果からOsCERK1は微生物に対する共生と排除という正反対の二重機能を担っており、おそらくは受容体のパートナーを換えることで応答を切り替えていることが推測されました。イネ以外の多くの植物でも、CERK1に共生応答を起動する能力があるのは以前に示した通りです。CERK1がMycファクターを受容しているかどうかまでは証明していませんが、NFR1との類似性を考えると可能性は高いでしょう。この時点で私は、この論文が相当なインパクトを持つことを確信しましたが・・・・
コメント