
Stougaard博士のNIN遺伝子の単離報告と前後して、ミヤコグサ(Lotus japonicus)やタルウマゴヤシ(Medicago truncatula)から、根粒菌共生が破綻した変異体が多数報告されました。その中でも歴史的な経緯から、Nodファクター受容体が特に注目されていて、誰が一番最初に単離するか?という競争になっていました。ちなみにNINは転写因子をコードしており、表現型もNodファクター受容体とは考えられません。
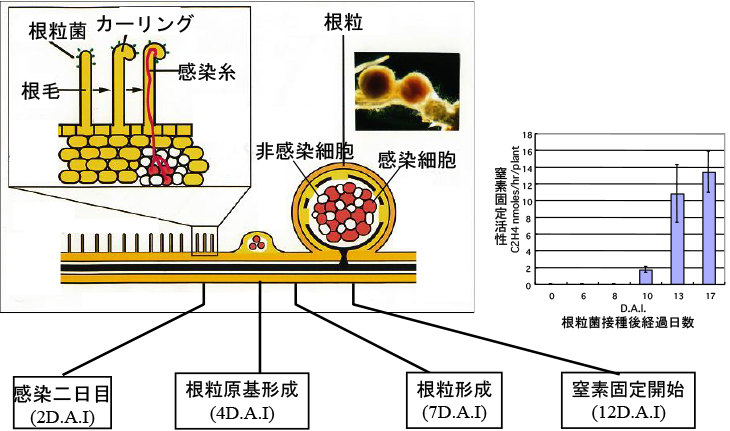
変異体の原因となった遺伝子の同定方法については、いずれどこかで解説したいと思うのですが、当時は非常に時間と手間がかかりました。単離された変異体の中にNodファクターの受容体の変異体が含まれていれば、全ての共生応答が停止しているはずです。その頃に私が知っていた最も早い共生応答は根毛のカーリングですが、これは感染から3-4日しなければ見えません。カーリングしない変異体は沢山あるので、もっと早い共生応答を見つけて整理する必要があります。おそらくヨーロッパやアメリカの共生研究者達は、この辺りの展開も予想して準備していた・・・というのは買いかぶり過ぎだと思いますが、共生分野の苦難の90年代にはそのような解析も行われていました。
数々の生理学実験から、共生で本当に意味があるかはともかくとして、カーリング以前のイベントが報告されています。Nodファクターを投与して1分以内には根毛の先端でカルシウムやカリウムなどのイオンの流入や排出が生じて、さらに細胞質のpHが変動します。15分後には根毛細胞の核周辺のカルシウム濃度が増減を繰り返す“カルシウムスパイキング”が生じます。数時間後には根毛の先端が変形したり分岐したりします。
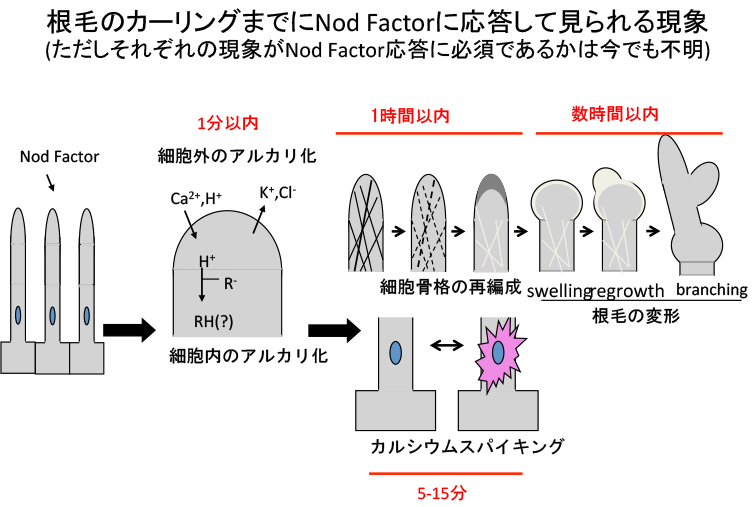
このような指標を元に単離された変異体を分類してみると、カルシウムスパイキングやイオン流入の有無が変異体によって異なることが判りました。この情報を基に時系列に従って変異体を並べると、下図のようになります。
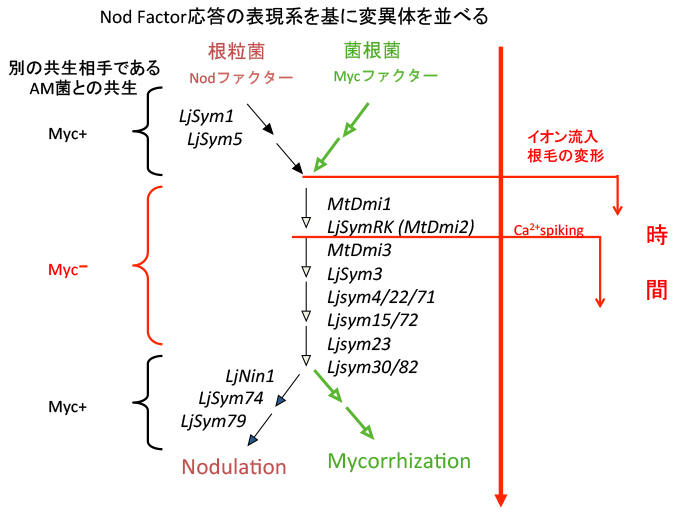
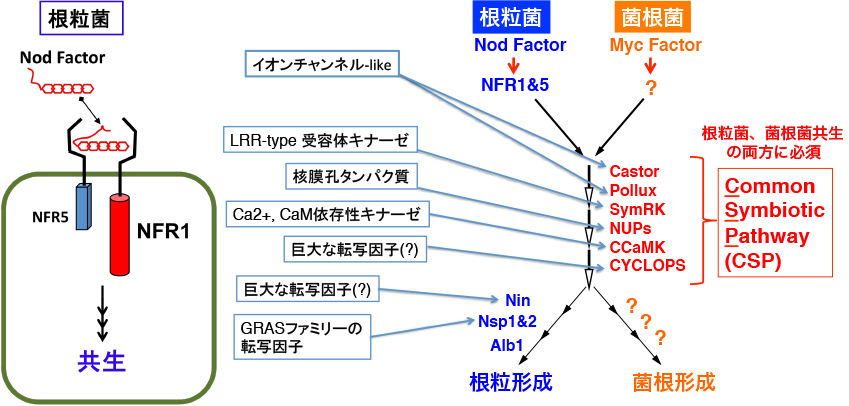
Ljsym1やLjsym5は、このようなNodファクター応答も全て破綻しています。またLjSymRKはイオン流入は生じますが、カルシウムスパイキングは生じません。この時に海外の研究者達は、面白い事実に気が付きました。根粒菌とは全く異なる微生物で、陸上植物の大部分と共生するアーバスキュラー菌根菌(AM菌。別のページで詳しく解説します)という微生物の感染が、これらの変異体の一部では破綻しているのです。面白いことにLjsym1やLjsym5はAM菌共生は破綻していません。さすがにAM菌の共生シグナルが、Nodファクターと同一であるとは考えにくいので、Ljsym1やLjsym5がNodファクター受容体の変異体であるという期待が高まります。そして2003年にStougaard博士らによって、これらの変異体の原因遺伝子が細胞膜に局在するLysM型受容体キナーゼというタンパク質をコードしていることが報告されました(2003年のNature, 424, 637-640とNature, 425, 585-592)。ちなみにLjsym1やLjsym5という名前は変異体を取る時の管理名で、原因遺伝子はそれぞれLjNFR1, LjNFR5と名付けられています。これと前後して、他の変異体の原因遺伝子も続々と同定されて行きました。
コメント