

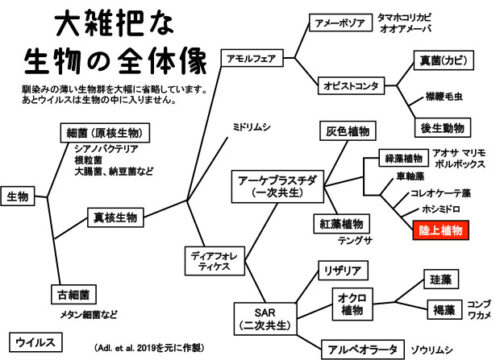
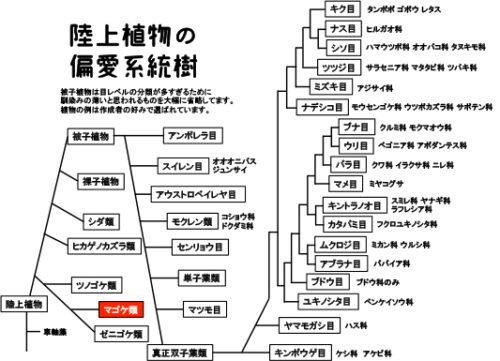
真核生物>ディアフォレティケス>アーケプラスチダ>ストレプト植物>コケ植物>マゴケ類(蘚類)>ヒョウタンゴケ目>ヒョウタンゴケ科>ニセツリガネゴケ属>ヒメツリガネゴケ
コケ植物はツノゴケ類、マゴケ類(蘚類)、ゼニゴケ類(苔類)の3つに大別されます。研究分野で圧倒的に有名なコケ植物がヒメツリガネゴケ(蘚類)とゼニゴケ(苔類)です。どちらもコケ植物の研究だけではなく、植物科学全般の理解に大きく貢献しています。
例えば花について、モデル植物としてもっとも有名なシロイヌナズナやイネ(被子植物)で解析が進んでいますが、あまりに多くの遺伝子が複雑に花の形態や咲く時期を制御しています。これらの複雑な仕組みがいきなり花を形成し始めた裸子植物で獲得された・・・訳では無く、どうやらシダやコケ植物の段階で既に基本的な仕組みが存在しており(生殖器形成などに使われている)、それが転用されて花が獲得されたことが判ってきました。もちろん裸子植物や被子植物になってから付け足された仕組みも多いでしょうが、コケ植物から原始の仕組みを類推できるメリットは非常に大きいのです(今までは足し算や引き算もわからないのに微分積分を解かなければならなかったようなもんです)。もちろん花だけではなく、他の様々な器官の形態形成や病原抵抗性、光応答など非常に多くの分野にコケ植物が貢献しています。
このようなコケ植物自体の重要な進化的立場に加えて、ヒメツリガネゴケには他の植物の研究者から見ると「反則だ!!」と言いたくなるほどの利点がありました。遺伝子レベルの仕組みを解明するためには、注目した遺伝子を破壊して影響を調べることが重要です。マウスや酵母などでは、早くから「相同組換え」という仕組みによる狙った遺伝子の破壊が可能でしたが、植物分野ではもっとも整備されたシロイヌナズナでさえも、最近まで不可能でした(CRISPR-Cas9・・・ありがとう!!)。このために研究者は多大な労力をかけて回り道をしていたのです。ところがヒメツリガネゴケは、例外的に早くから狙った遺伝子を壊す方法が確立されていました。また被子植物は2倍体以上なので、一方の染色体の注目する遺伝子が壊れても、もう一方が相補してしまって影響が観察できないことが多いのです。そこで自家受精(不可能な植物もある)させて、孫の代から両方の染色体の遺伝子が壊れたホモ個体というのを選抜する必要があるのですが、生活環の大部分が半数体(1倍体)のコケ植物では、このような手間もありません。私はシロイヌナズナと比較しても圧倒的に使い難いマメ科植物の研究者ですが、コケ植物の利便性を知った時には卒倒しそうになりました。
ヒメツリガネゴケの研究における利便性は、基礎生物学研究所の紹介動画も参考にしてください。とても綺麗で面白いです。
研究材料として以外のヒメツリガネゴケの魅力ですが・・・・・、まあコケ植物全般に言えることかもしれませんが、再生能力が非常に高いことです。あと胞子体がかわいい!! 以上です(わかり次第書き足しますが・・・・・・・・・・・・・・・・・・・・・・・あるのかな?)。
コメント