
くわしくは・・・こちら
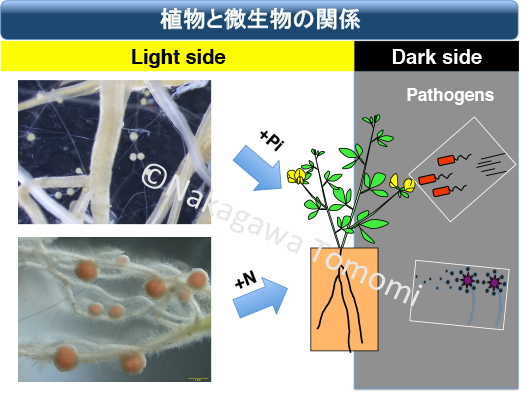
マメ科植物は病原菌を排除しながらも根粒菌を受け入れます。病原菌は進化が早く、さらに巧妙な感染手段を色々と備えているのですが、マメ科植物はどのようにして病原菌を排除と根粒菌共生を両立させているのでしょうか?
根粒菌が分泌する共生シグナルのNodファクターは、カビの細胞壁成分で防御応答を誘導することが知られているキチンを基本骨格としています。またマメ科植物のNodファクターを認識する受容体NFR1は、キチンを認識して防御応答を起動するCERK1と、非常に類似しています。私はこれを手がかりに防御と共生の関係を調べました。
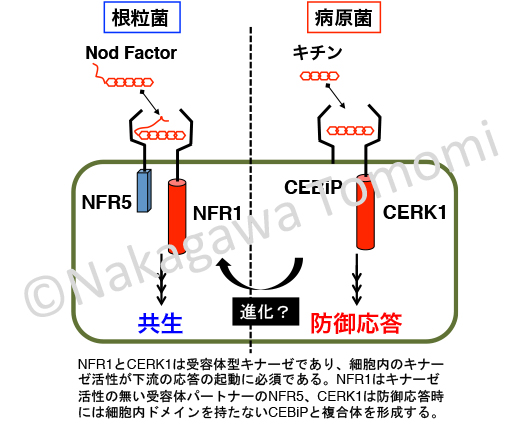
防御応答の起動しかできないシロイヌナズナのAtCERK1と、ミヤコグサのNFR1の細胞内ドメインを比較解析したところ、共生応答はNFR1キナーゼドメインの3アミノ酸のYAQ配列が重要な役割を果たしていることを突き止めました。

シロイヌナズナは、陸上植物の中でも例外的にアーバスキュラー菌根菌(AM菌)と共生しないアブラナ科に属します。意外なことにAM菌共生するイネやトマトなどの多くの非マメ科植物のCERK1は、キナーゼドメインにYAQや類似したYAR配列を保持していました。しかもこのキナーゼドメインは、ミヤコグサでNFR1のキナーゼの代わりに根粒菌共生を起動できます。そこでCERK1がAM菌共生に関与している可能性を調べるために、イネでCERK1が壊れた変異体を作出して解析したところ、なんとイネのoscerk1変異体は防御応答もAM菌共生も破綻していました。つまりOsCERK1は、単独で病原菌の排除と共生菌の受け入れという正反対の生理応答の起動に関わることが明らかとなったのです。

くわしくは・・・こちら
コメント