
詳しくは・・・こちら
自然界で植物は様々な病原菌に狙われており、それに対抗して複雑な防御メカニズムを展開しています。たとえば糸状菌(カビ)は10万種以上知られていますが、この中でイネに加害できるのは約50種に過ぎず、さらに深刻な被害を起こせるのは10種以下だと言われています。つまり植物は大多数の病原菌の侵攻を阻止することができますし、その一方で高度な防御メカニズムを突破できる病原菌も存在しています。植物と病原菌は、永い進化の過程で対抗進化を繰り返しながら、お互いの存在を賭けて戦っているのです。
このような視点で見た場合に、宿主植物が微生物を受け入れて相利共生を行うことは奇異に感じられます。実際に私は根粒菌共生において、根粒菌を受け入れるプログラムを発動させるNFR1が、防御応答を起動させるCERK1から進化した可能性が高いことを示しました(Nakagawa et al., 2011; 植物の敵と味方 〜承〜)。また陸上植物の大部分(マメ科含む)が受け入れるアーバスキュラー菌根菌(AM菌)共生では、CERK1自体が共生の開始に重要な働きをしていることを明らかにしました (Miyata et al., 2014; 植物の敵と味方 〜転〜 3)。NFR1も防御応答の起動能力自体は保持しており(植物の敵と味方 〜起〜1)、しかもAM菌共生を行う被子植物のCERK1は防御と共生の二重機能を保持しているようです。
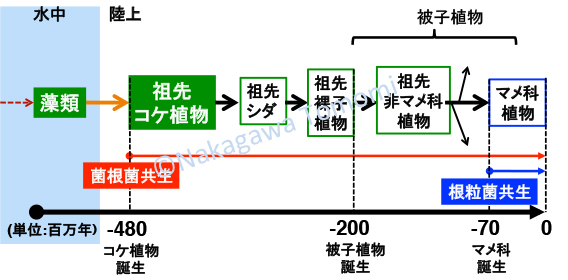
永い進化の過程では、多くの遺伝子が重複して機能分化することで、維管束や花・種子などの多くの形質が獲得されていきました。AM菌共生は最初の陸上植物であるコケで成立したと考えられており、不便に見える防御と共生の二重機能を、それぞれ「防御」と「共生」に分けて専門化&最適化する時間は充分あったはずです。ただし、原始の共生で本当にCERK1の二重機能が保持されているか不明であるので、私はコケ植物のCERK1が防御と共生の二重機能を持っているかを調べることにしました。

既にモデルとして確立されているコケ植物であるヒメツリガネゴケやゼニゴケは、残念ながらAM菌と共生しない例外的な植物です。そこで私達は、AM菌共生を行うことが知られてるフタバネゼニゴケ(Marchantia paleacea subs.diptera)をモデルとして整備し始めました。フタバネゼニゴケがキチン防御応答を起動できることや、AM菌共生することも確認して、次世代シーケンサーによるRNA-seqで共生や防御に関わる遺伝子の大部分が保持されていることを確認しました。またCERK1も1コピー存在しており、共生応答の起動に必要なYAQ/YAR配列が保存されていました。さらにフタバネゼニゴケのCERK1のキナーゼドメインが、マメ科植物でNFR1の代わりに共生プログラムを起動できることも確認できました。一方でモデルのゼニゴケとは異なり、フタバネゼニゴケに胞子を形成させることができないために、CERK1を破壊した株を作製して、本当にCERK1が防御と共生に関わるかどうかを証明できていません。しかしフタバネゼニゴケでもCERK1の二重機能は保持されている可能性が高いです。
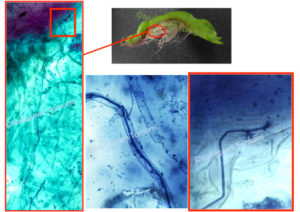
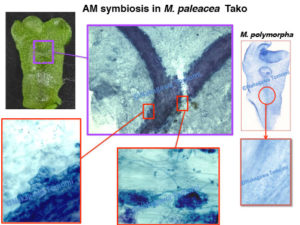
根粒菌やAM菌は宿主植物に受け入れてもらうために、NodファクターやMycファクターなどの身分証を提示する必要があります。受け入れられてしまえば、宿主植物から至れり尽くせりのおもてなしを受けることができます。これは病原菌にとっては非常に羨ましいことでしょう。
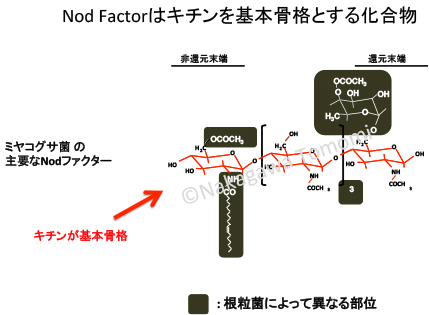
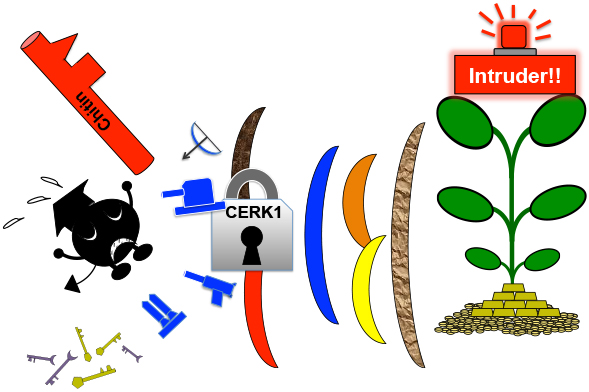
植物病理学分野を見ていると、病原菌は信じられないくらい巧妙な手段を次々と開発して感染しようとします。したがってNodファクターやMycファクターを偽造して侵入を試みるのは当然の戦略のハズですが・・・、実際には共生経路を悪用して侵入する「病原菌」は知られていません。つまり植物にも相応の対抗策が備えられているはずです。
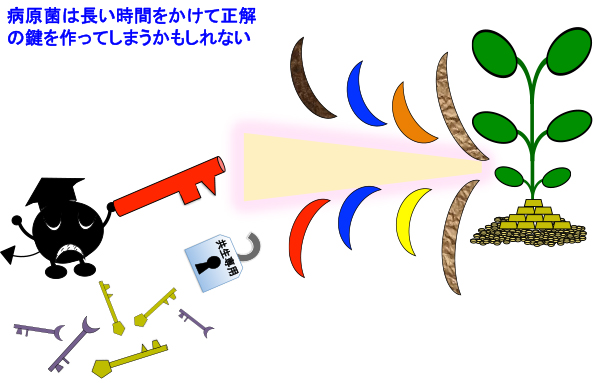
仮に共生専用のNFR1やCERK1を作ってしまうと、ペナルティさえ無ければ延々と試すことで鍵を開けることは可能でしょう。4〜5億年の時間というのは、特に進化の早い微生物にとっては充分過ぎる時間です。
しかし共生の鍵が防御応答を発動させるキチンを少し改変したもので、さらに錠が防御応答の発動装置を兼ね備えており、間違うと防御応答が発動するのであれば、相当に難易度が上がると推測されます。4〜5億年の時間を耐えるほどの工夫であるとは思えないですが、NFR1やCERK1が二重機能を維持しているのはコレが理由ではないでしょうか・・・・????
コメント