

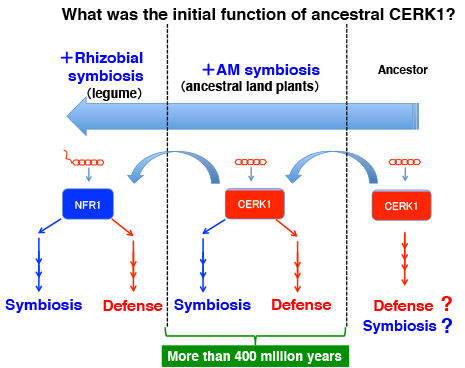
フタバネゼニゴケは、期待したとおりAM菌共生とキチン防御応答の能力を備えていました。それではCERK1を持っているのでしょうか?
当然ながら誰もゲノム配列を調べたことが無い生物なので、自分で明らかにするしかありません。そこで東京農工大学におられた古崎利紀博士らのご協力で、次世代シーケンスによるトランスクリプトーム解析を行いました。
その結果、意外なことに被子植物で知られていたAM菌共生に必要な遺伝子の大部分が、フタバネゼニゴケにも保存されていることが判明しました。もちろんCERK1もです。しかし面白いことに、マメ科植物でNFR1のパートナーとして根粒菌共生シグナルの受容に必須のNFR5が、フタバネゼニゴケでは見つかりません。2011年当時、我々はCERK1が本当にAM菌共生に関わっているかどうか証明出来ていませんでしたが、ほぼ確信していました。もちろんCERK1は防御応答を起動する受容体なので、共生応答を起動するにはシグナルを切り替える何かが必要です。
NFR1の非マメ科植物の相同遺伝子はCERK1ですが、NFR5は少なくとも非マメ科の被子植物にも保存されています。私達はそのNFR5が防御応答から共生応答に切り替える鍵だと予想していたのですが・・・・(その後に我々は、少なくともイネのNFR5が共生に関わっていないことを示しました。植物の敵と味方 〜その後〜)。
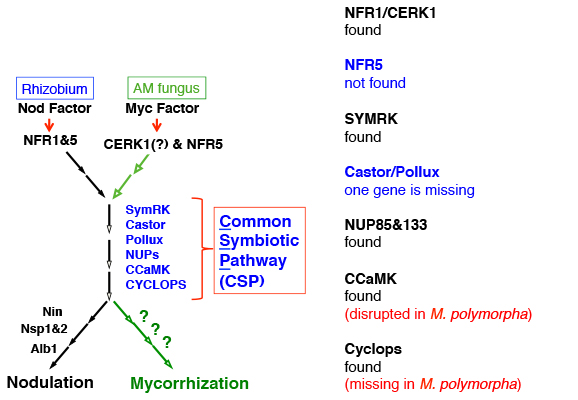
また面白いことに、AM菌共生を行わない例外的な植物であるゼニゴケ(Marchantia polymorpha)のゲノム情報を京都大学の河内孝之博士に提供していただき、古崎さんらに解析していただいたところ、なんと共生に必須のCCaMKとCyclopsが壊れていることが判明しました。当時は河内研に所属されていた石崎公庸博士が、ゼニゴケにフタバネゼニゴケのCCaMKやCyclopsを導入して下さったのですが、AM菌共生の機能は戻らないようです。
ちなみにこの時に被子植物で知られている防御応答遺伝子も調べたのですが、やっぱりゼニゴケでは冗長性が著しく低いことが判りました。

被子植物で知られている防御遺伝子でゼニゴケで見つからないのは、私が少し調べた範囲では細菌の鞭毛成分を感知するFLS2くらいです。活性酸素を発生させるRboHに似た遺伝子などは、シロイヌナズナでは途中で書き出すことを挫折するくらい見つかります。しかしフタバネゼニゴケやゼニゴケでは2つで、しかも防御応答を担う分子種と形態形成(根毛の先端成長などで活性酸素が関わる)に関わる分子種ではないかと推測されます。全く被子植物と似ていなければ使う意義が薄いですが・・・・、似たシステムを持っている上で単純というのは非常に強力です。
コメント