
フタバネゼニゴケに胞子を形成させることができません。そして胞子が無いと、フタバネゼニゴケが持っているCERK1を破壊することができません。このプロジェクトのゴールが「コケのCERK1に防御と共生の二重機能が保持されているか?」なので、CERK1を破壊して確認できないのは大問題です。
結論から言いますと、現時点でもフタバネゼニゴケのCERK1破壊株は作製出来ていません・・・。色々と工夫して、遺伝子導入ができるようにはなっており、共同研究者は他の遺伝子の破壊に成功し始めているのですが。

CERK1や他のAM菌共生遺伝子群は、陸上植物と進化的に近縁の藻類にも多くが保存されていますが、少し遠縁になると保存されていません。またフタバネゼニゴケでは、遺伝子破壊株による完全な証明はできていませんが、防御や共生に関与していることが示唆されています (防御と共生の進化 4、防御と共生の進化 6)。どうやらコケ植物よりも前の段階が、「CERK1はどちらの機能を先に獲得されたのか」に答えるための重要なポイントのようです。
つまりCERK1はコケでも防御と共生の二重機能を保持している可能性が高い。私達はイネでCERK1の二重機能を証明しましたが、イネとコケは4億年以上前に共通祖先から分岐したと考えられます。つまりCERK1の二重機能は、4億年以上の陸上植物進化の年月で程存されていたことが推測されます。
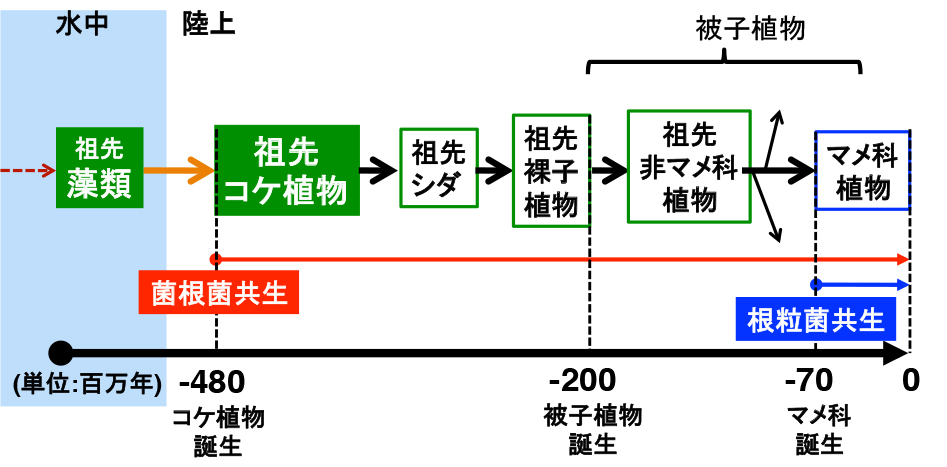
祖先コケ植物から被子植物が進化する過程で、維管束や花・種子の獲得などの目に付きやすい形質だけでも相当に変化しました。他にも多くの形質が進化したことでしょう。この原動力になるのが、遺伝子の「重複」と「機能分化」です。
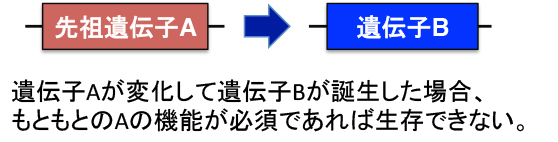
ある遺伝子から新しい遺伝子が生じる場合、もとの機能が重要であれば、その機能を失う訳にはいきません。このような変異が生じた子孫は生存競争から脱落するでしょう。
現在生き残っている生物を解析すると、変異前に遺伝子が重複して、一方が元の機能を果たしている間に他方が進化することが多いようです。
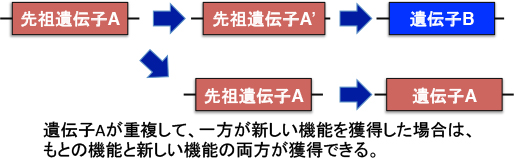
つまり何が言いたいかと言いますと、祖先コケ植物から現在の被子植物までの間に、防御と共生という正反対の二重機能を持つCERK1が重複して、「共生専用CERK1」と「防御専用CERK1」に分かれる時間は充分あったと考えられます。しかし実際には、大部分の植物でCERK1は1コピーしかありませんし、AM菌共生を行わないシロイヌナズナでは「防御専用CERK1」が見つかっていますが、「共生専用CERK1」を持つ植物は知られていません。また根粒菌共生の開始を担うマメ科植物のNFR1も、CERK1と同様に防御応答を起動する能力を維持したままです (植物の敵と味方 概要)。
「共生専用CERK1」を産み出すことは難しく無いと想像されますので、おそらくは・・・全て生存競争から脱落するほどの悪手なのでしょう。
その理由として私が考えているのが、違法アクセスに対するペナルティという仕組みです。
マメ科植物のNFR1は、「アクセスしてきた根粒菌を受け入れるか否か?」という判断をする重要な遺伝子です。そして、おそらくCERK1も「アクセスしてきたAM菌を受け入れるか否か?」の選択を担っている可能性が高い。
受け入れられた共生菌は、宿主植物から至れり尽くせりのおもてなしを受けるのですが(至れり尽くせりの根粒菌? その2)、これは他の微生物にとっては魅力的でしょう。私が病原菌であれば、NFR1やCERK1を誤魔化して受け入れプログラムを作動させることを考えます。
(病原菌と植物の攻防はこちらを参照)
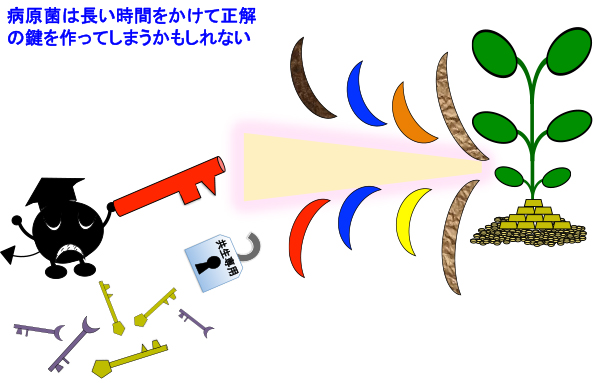
しかし共生を起動するための鍵が防御応答を起動させるキチンに類似した物質であり(植物の敵と味方 〜序〜 4)、それを認識するNFR1やCERK1が防御応答の起動装置も兼ねていれば、不正にアクセスする難易度が上がると推測されます。CERKやNFR1の二重機能性は、これが理由であると私は考えています。
面白いことに、根粒菌共生やAM菌共生の仕組みを悪用して不正に侵入する病原菌は、今まで全く知られていません。植物の防御応答に常に悩まされている病原菌にとっては、積極的に植物が受け入れてくれる共生のシステムは非常に魅力的だと思うのですが・・・。不正アクセスを試みない訳が無いと私は思うのですが、おそらく不正アクセスを許してしまうような間抜けな宿主植物は、全て進化の過程で脱落しているのでしょう。
コメント